
LSD, Entropy and Meta-State Transitions
Open-source LSD fMRI data reveals a higher rate of meta-state transitions in the psychedelics state

Introduction
That we are amidst a Psychedelic Revival has become obvious, such that it is impossible to begin a paper on psychedelics without an adroit acknowledgement of this fact. The extent to which these 'mind-manifesting' compounds have captured the public imagination is perhaps best exemplified by the valuation of the psychedelic behemoth COMPASS PATHWAYS at over 1 Billion USD. [Ettinger, 2021]
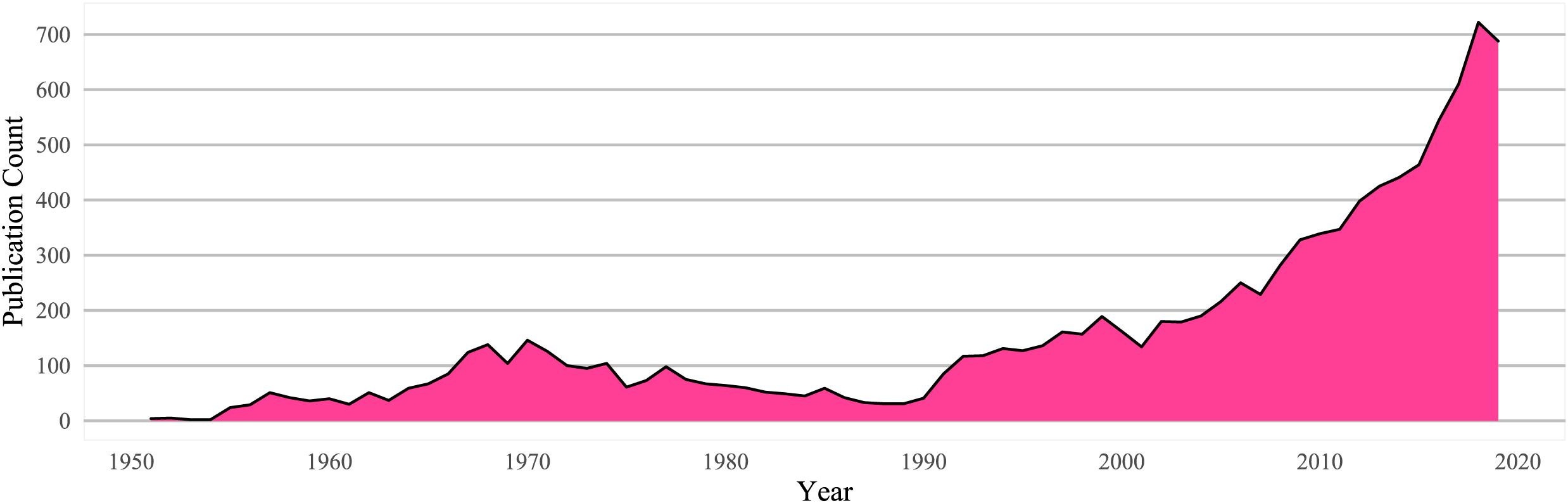
While there is ample evidence that the use of psychedelia long predates the modern era, especially by indigenous groups in Africa, Asia and the Americas, they entered into the mainstream only in the 1950's, post the discovery of LSD by Albert Hoffman in 1943 and Gordon Wasson's introduction to psilocybin mushrooms through Maria Sabina in 1955. Scientific research into the effects of psychedelics blossomed in the 60's and early 70's before being abruptly ground to a halt due to the whiplash of regulations following the tumult of the counterculture. Research slowly began to pick up in the 90's following the pioneering work by Rick Strassman with DMT and continued to pick up in the 2000's led by researchers at John Hopkins, with Roland Griffiths and colleagues' seminal paper 'Psilocybin can occasion mystical-type experiences having substantial and sustained personal meaning and spiritual significance' in 2006 [Griffiths, 2006] being a watershed movement, evidenced by the paper being cited over a thousand times. The exciting beginnings, dramatic fall and subsequent enthusiastic comeback of psychedelic research is captured nicely by the publication count by year in Figure 1
📌 Figure 1 [Petranker, 2020] -
By definition, all classic psychedelic drugs are psychoactive compounds that primarily produce their effects via agonism at serotonin 2A (5-HT2AR) receptors [Glennon, 1984]. The canonical list of psychedelics thus are LSD (Lysergic acid diethyl-amide), psilocybin (active component of "magic mushrooms”), DMT (N,N-Dimethyltryptamine) and mescaline (found in Peyote and San Pedro cacti). While the compounds in question may have remained (for the most part) the same, the study of psychedelics has markedly changed since the 60's. The most pertinent difference is the availability of sophisticated brain-imaging methods such as functional Magnetic Resonance Imaging (fMRI) and Magnetoencephalography (MEG) that allow for a much deeper exploration into the physical mechanisms of psychedelic action in the brain.
fMRI has proven to be an attractive option for psychedelic researchers, in particular, resting-state fMRI (rsFMRI) where-in participants simply lie in the scanner without any tasks or stimuli. By comparing rsFMRI data during drug effects and placebo, one can purport to investigate the fundamental changes in measures of whole-brain network dynamics such as resting-state functional connectivity(RSFC) during a psychedelic experience. This data can then be further related to behavioural measures.
The first rsfMRI study was conducted by Robin Carhart-Harris and colleagues at Imperial College London in 2012 with 15 participants using psilocybin. Since then, according to a meta-review performed by McCulloch and colleagues in 2021, 37 psychedelic rsfMRI papers have been published based off data from 16 unique rsfMRI datasets. A description of these datasets, along with the timeline of publications, is provided below -
📌 Figure 2 [McCulloch, 2021] -

Based off their meta-analysis, the authors made the important observation that "no two studies employ the same data processing and analysis strategy. Two datasets are the foundation of more than half of the published literature". The two foundational datasets mentioned are the Carhart-Harris 2012 and 2016 datasets with psilocybin and LSD respectively and are the only two datasets, in addition to the Palhano-Fontes 2015 Ayahuasca dataset, that are currently open source.
The authors further note that "of the 37 articles, 15 apply a “seed-based connectivity” analysis wherein the time series from a predefined region (or regions) is correlated against the time series from all voxels in the brain" and a further "twelve articles employed a network-based connectivity approach". Finally, they note that 6 of the studies investigated changes in the entropy of the RSFC, although each study calculated entropy using different methods such as Lempel-Ziv [Varley, 2020] and Shannon entropy [Lebedev, 2015]. The authors conclude their paper with a call towards orienting around open science principles and stressing the need to avoid the pitfalls that lead to the replication crisis. Their primary suggestion was to align methodologies, such as those used to calculate entropy, in a commensurable manner.
The goal of this paper will be to propose a potential method for doing so via the adoption of an intuitive, robust and standardised measure of resting-state brain activity — the rate of meta-state transitions (MSTs). The methodology and empirical underpinnings of MSTs comes from Tseng and Poppenk's 2020 Nature Communications paper 'Brain meta-state transitions demarcate thoughts across task contexts exposing the mental noise of trait neuroticism'. [Tseng and Poppenk, 2020] ****This methodology will be applied to the open-source Carhart-Harris 2016 LSD data with the hypothesis that the rate of meta-state transitions will be higher in participants under LSD as compared to placebo.
Background
Entropy
Before proceeding with the rest of the paper, it will be useful to lay out some theoretical background. A reigning theory for explaining the effects of psychedelics is the 'Entropic Brain Hypothesis' proposed by Robin Carhart-Harris and its follow-up REBUS (Relaxed Beliefs Under Psychedelics) co-proposed with Karl Friston. The entropic brain hypothesis posits that “the defining feature of [psychedelic states] is elevated entropy in certain aspects of brain function, such as the repertoire of functional connectivity motifs that form and fragment across time.” Here, entropy is construed as a “dimensionless quantity that is used for measuring uncertainty about the state of a system“ and can be thought of as non parametric measure of variance. The theory also makes the claim that the richness of subjective experience, or qualia, is indexed by the level of entropy in the brain and thus offers a bridge between the subjective-objective realms. [Carhart-Harris, 2014]
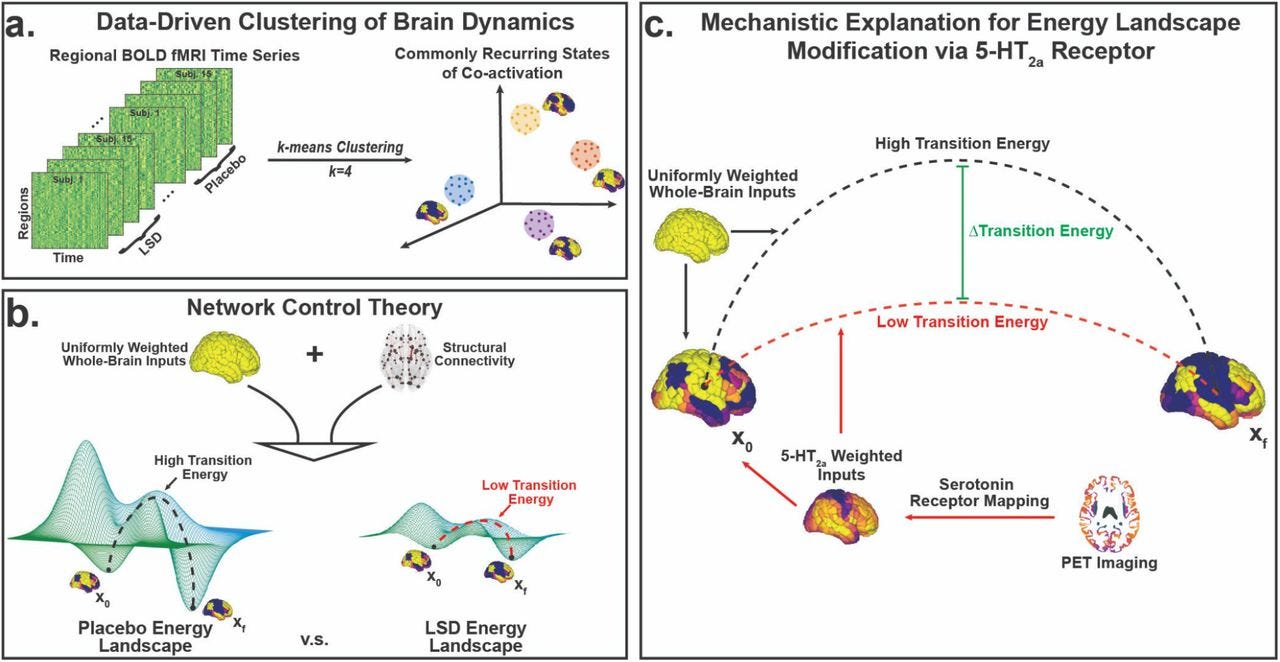
REBUS extends the entropic brain hypothesis by combining it with Friston's Free Energy Principle to make the claim that "via their entropic effect on spontaneous cortical activity— psychedelics work to relax the precision of high-level priors or beliefs, thereby liberating bottom-up information flow, particularly via intrinsic sources such as the limbic system." [Carhart-Harris & Friston, 2019] In other words, psychedelics flatten the free-energy landscape of the brain. Empirical evidence for the theory was provided recently by Singleton and colleguages who combined PET and fMRI data with Network Control Theory to show that serotonin 5HT2A agonism lead to a lower required transition energy between certain recurring brain states.
📌 Figure 3 [Singleton, 2021] -
Meta-State Transitions
“When we take a general view of the wonderful stream of our consciousness, what strikes us first is the different pace of its parts. Like a bird's life, it seems to be an alternation of flights and perchings. The rhythm of language expresses this, where every thought is expressed in a sentence, and every sentence closed by a period." - William James, Stream of Consciousness [James, 1892]
Investigating the nature of thoughts is a primary concern for the brain and behavioral sciences and much research has been devoted to teasing apart the contents of our inner sanctum. However, it is also insightful to consider not just the 'what' of thoughts but also the 'how'. As James elegantly notes, our stream of consciousness seems to proceed in a sequence of flights and perchings, i.e. – thoughts and transitions. What would be the nature of these transitions and can we use fMRI data to identify them in a coherent way?
This is the question that animates Tseng and Poppenk's 2020 Nature Communications paper [Tseng & Poppenk, 2020]. They propose adopting a “constructionist model of the mind", that is, "complex mental states emerge from flexible network-level interactions, and changes in active network configurations (i.e., time-varying functional connectivity) might signal boundaries between cognitive states”. These changes in active network configurations, or meta-state transitions, can be thought of as demarcations of thought.
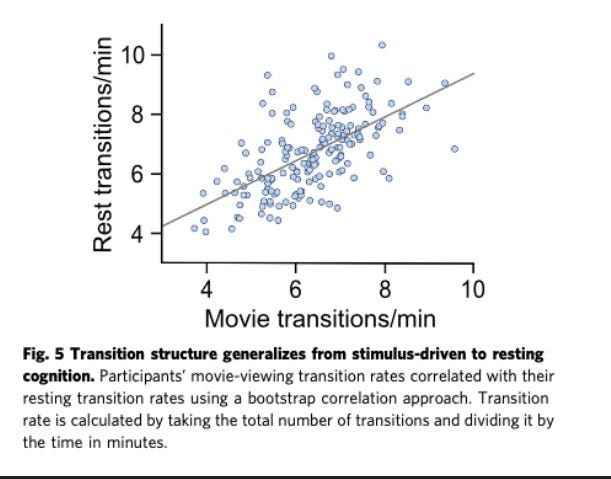
In their paper, they identify meta-state transitions for both movie watching fMRI data and resting-state fMRI data for the same subjects . They found that "meta-state transitions during movie-viewing best predicted event boundaries and onset of new semantic information support the interpretation that meta-state transitions align with the changes in semantic content across one’s thoughts”. Their calculated transition rate for the movie watching data was highly consistent across runs, was stable across tasks and days and was correlated with the transition state at rest. Finally, higher levels of trait neuroticism were associated with higher resting transition rates, implying trait-level characteristics could be deduced from such data.
📌 Figure 4 [Tseng and Poppenk, 2020] -
Data
LSD
The LSD data was first published and described in the 2016 PNAS paper 'Neural correlates of the LSD experience revealed by multimodal neuroimaging' by Robin Carhart-Harris and colleagues at Imperial College London [Carhart-Harris, 2016]. A total of 20 subjects were recruited by word of mouth with the key exclusion criteria including being less than "21 years of age, personal history of diagnosed psychiatric illness, immediate family history of a psychotic disorder, an absence of previous experience with a classic psychedelic drug, any psychedelic drug use within 6 weeks of the first scanning day, pregnancy, problematic alcohol use, or a medically significant condition rendering the volunteer unsuitable for the study." Participants attended two study days that were separated by at least 14 days, receiving placebo on one and LSD on the other, with the conditions balanced across participants and the participants (but not the researchers) blind to the order.
Imaging was performed on a 3T GE HDx system. The fMRI data consists of 3 fMRI runs of 7.20mins (440secs) each. All the runs were closed-eye no-task with the participants encouraged to close their eyes and relax in a reclined position when the drug was administered. One of the runs (Run 2) involved participants listening to music. All participants received 75 µg of LSD, administered intravenously via a 10ml solution. MRI scanning started approximately 70 minutes post dosing, and lasted for approximately 60 minutes (this included the structural MRI scan).
BOLD-weighted fMRI were acquire with the following parameters - "a gradient echo planer imaging sequence, TR/TE = 2000/35ms, field-of-view = 220mm, 64 × 64 acquisition matrix, parallel acceleration factor = 2, 90° flip angle. Thirty five oblique axial slices were acquired in an interleaved fashion, each 3.4mm thick with zero slice gap (3.4mm isotropic voxels)"
One subject did not complete the bold scans and four other subject's scans were discarded due to excessive head movement, hence a total of 15 subjects remained. This data was preprocessed with the following protocol [taken from the Supplementary Appendix of Carhart-Harris, 2016] -
removal of the first three volumes;
de-spiking (3dDespike, AFNI);
slice time correction (3dTshift, AFNI);
motion correction (3dvolreg, AFNI) by registering each volume to the volume most similar, in the least squares sense, to all others (in-house code);
brain extraction (BET, FSL);
rigid body registration to anatomical scans (twelve subjects with FSL’s BBR, one subject with Freesurfer’s bbregister and two subjects manually);
non-linear registration to 2mm MNI brain (Symmetric Normalization (SyN), ANTS);
scrubbing (8) - using an FD threshold of 0.4
spatial smoothing (FWHM) of 6mm (3dBlurInMask, AFNI);
band-pass filtering between 0.01 to 0.08 Hz (3dFourier, AFNI);
linear and quadratic de-trending (3dDetrend, AFNI);
regressing out 9 nuisance regressors : out of these, 6 were motion-related (3 translations, 3 rotations) and 3 were anatomically-related (not smoothed). Specifically, the anatomical nuisance regressors were: 1) ventricles (Freesurfer, eroded in 2mm space), 2) draining veins (DV) (FSL’s CSF minus Freesurfer’s Ventricles, eroded in 1mm space) and 3) local white matter (WM) (FSL’s WM minus Freesurfer’s subcortical grey matter (GM) structures, eroded in 2mm space).
Human Connectome Project
To generate the spatial maps needed for dual regression, the 2017 Human Connectome Project Young Adult data was utilised. Specifically, the group-ICA data from 1003 subjects who completed four compete rsfMRI runs was used, as described in the 'HCP1200 July 2017 release of high-level rfMRI connectivity analyses'. The details of the subject recruitment and data acquisition can be found in the 2013 NeuroImage paper 'The WU-Minn Human Connectome Project: An overview'. [Van Essen, 2013] Briefly, subjects were drawn from the a population of twins in the age range 22-35 and two 2hr scans were performed on a customised 3T scanner at Washington University with a multiband factor of 8, spatial resolution of 2 mm isotropic voxels, and a TR of 0.7 s. Out of the 2hrs, the first 30mins were two rsfMRI scans of 15mins each. The processing of the data to generate the group-ICA data was described as follows -
"Each 15-minute run of each subject’s rfMRI data was preprocessed according to [Smith 2013]; it was minimally-preprocessed [Glasser 2013], and had artefacts removed using ICA+FIX [Salimi-Khorshidi 2014, Griffanti 2014]. Inter-subject registration of cerebral cortex was carried out using areal- feature-based alignment and the Multimodal Surface Matching algorithm (‘MSMAll’) [Robinson 2014, Glasser 2016]. For feeding into group-PCA, each dataset was then temporally demeaned and had variance normalisation applied according to [Beckmann 2004]. Group-PCA output was generated by MIGP (MELODIC’s Incremental Group-PCA). This output comprises the top 4500 weighted spatial eigenvectors (eigenmaps) from a group-averaged PCA [Smith NeuroImage 2014].The MIGP group-PCA output was fed into group-ICA using FSL's MELODIC tool [Hyvärinen 1999, Beckmann 2004], applying spatial-ICA at several different dimensionalities (15, 25, 50, 100, 200, 300)."
Analysis
The LSD data was downloaded from OpenNeuro. A Python coding environment was created on Jupyter and nipipe was used to incorporate FSL tools. The Brain Imaging Data Strucutre (BIDS) and nibabel was used to access and load the data.
The workflow of the analysis is as follows (from Tseng & Poppenk, 2020]
📌 Figure 5 -

As described prior, the Human Connectome Project data has spatial maps of diffferent dimensionalities (15, 25, 50, 100, 200, 300). However, to be useful for the paper, the spatial maps had to be resampled to match the resolution of the LSD data, i.e changed from 2mm to 3.4mm voxel size. FSL's Melodic was used to resample the 15-ICA HCP data to produce 10 3.4mm spatial maps.
Next, FSL's dual regression was used to generate a time series for each spatial map (10xTR) for each subject's run. The time series were in the form of txt files with 10 columns for each spatial map time-series.
Then, the unsupervised, non-linear technique t-Distributed Stochastic Neighbor Embedding (t-SNE) was used to project the high-dimensional (10x) space to a low dimensional (2x) space. Scipy's manifold module was used with a perplexity of 30 and 1000 iterations for optimization. This resulted in each run consisting of a (2xTR) matrix. A visualization of this process from the Supplementary Appendix from Tseng and Poppenk's paper is provided below
📌 Figure 6 [Tseng & Poppenk, 2020] -

Next, the Mahalanobis distance was calculated for successive timepoints for each dimensionality reduced time-series to create a single step distance vector representing a measure for the similarity of brain network-activations, with larger distances indicating a greater dissimilarity between networks.
Peak distances in this step distance vector were considered meta-transition points and were selected using the peak algorithm of scipy.signal peak. A threshold value of 0.06 was used in accordance with Tseng and Poppenk's methodology. The total number of peaks thus identified in each run was stored.
Since the first 3 volumes were not included in the LSD data and the repetition time for this data was 2 seconds, the total fMRI scan was 434 seconds. To calculate the transition rate per min, the resulting number of peaks was divided by 434/60.
The above steps were performed 100 times for each run and the mean number of peaks (and thus the mean transition rate) was calculated.
Results
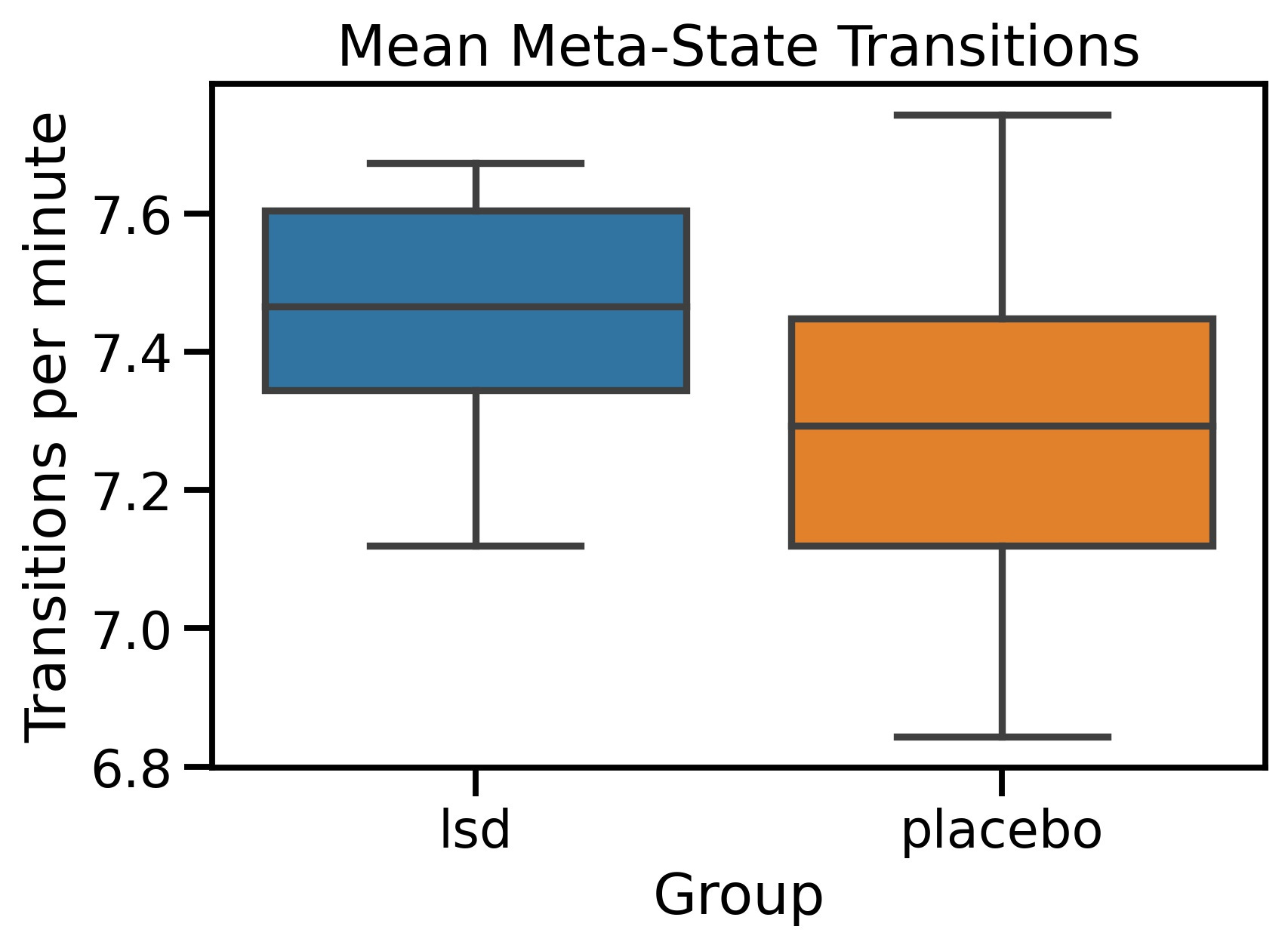
The mean meta-state transitions for each group are plotted below. The results lend support to my hypotheses that LSD would increase the number of meta-state transitions, although as the graph shows, my results are unlikely to be statistically significant. The Mean Transition rate for the LSD group was 7.47 (standard deviation 0.16) while for the placebo group it was 7.30 (standard deviation 0.24). A t-test was performed resulting in a t-statistic of 2.16 and a p-value of 0.04.
📌 Figure 7 -
Discussion
What does an increased meta-state transition rate mean? In the Tseng and Poppenk paper, the authors observe that an increased transition rate was positively correlated with higher scores of trait neuroticism, as measured by the Big5 questionnaire. Curiously, the authors also observed a negative correlation with trait openness, as shown below
📌 Figure 8 [Tseng & Poppenk, 2020] -

It is interesting to contrast this with the well-acknowledged and empirically backed observation that psychedelics lead to an increase in trait-openness. Indeed, here is Lebedev and colleagues noting in a 2016 paper that "Overall, LSD had a pronounced global effect on brain entropy, increasing it in both sensory and hierarchically higher networks across multiple time scales. These shifts predicted enduring increases in trait openness." [Lebedev, 2016] It should be noted that the increases in trait openness were observed two weeks after the LSD experiences. Perhaps, a higher trait-level transition rate leads to less openness while a transient increase in transition rate leads to more openness. Why this would be the case is open to interpretation.
Psychedelics increasing an individual's transition rate can be interpreted under the current paradigm as increasing the number of thoughts a person is having. But how does this jive with the actual psychedelic experience? For instance, the following figure, from the same 2016 LSD PNAS paper, displays the drastic changes in subjective effects precipitated by LSD
📌 Figure 9 [Carhart-Harris, 2016] -

We cannot really expect an increase in the number of thoughts per minute to lead to one experiencing a 'profound inner peace' or a 'sense of merging with my surroundings'. Thus, it it wise to avoid an overly simplistic interpretation of the transition rate that hastily attempts to cross the abyss of the hard problem of consciousness.
It was noted earlier that one of the motivations of this paper was to provide an intuitive, robust and standardised measure to probe psychedelic states. In a sense, such a measure already exists in the form of entropy. However, the lack of agreement on how best to calculate such entropy has lead to an inability to try and reproduce prior findings. While the relationship between meta-state transitions and entropy has not been explored in this paper, it seems likely that an increase in meta-state transitions is strongly correlated with increases in entropy by facilitating a greater ability to move between functional motifs. As meta-state transitions can be calculated without a priori assumptions, it could prove to be a useful tool to align various measures of entropy.
Finally, the observation of a correlation between trait-level states and the meta-transition rate suggests the possibility of the use of an individual's transition rate as a diagnostic tool. Alzheimer's and Parkinson's disease both intuitively seem to suggest a decrease or interruption in the flow of thoughts and thus could be detected earlier by a slowing of the transition rate. Most pertinently to the current moment, many individuals suffering from Long Covid exhibit a slowing of cognitive function and 'brain fog' that also suggests a potential decrease in transition rate. The potential of psychedelics as a treatment for such Long Covid symptomatology is an avenue worthy of exploration.
Limitations
A whole other paper can be written on the limitations of the various concepts and findings discussed in this paper, so this section will be focused on one broader methodological issue as well as particular limitations of my own results.
Resting-state functional connectivity is an often critiqued mode of inquiry as detractors argue fMRI data it is often too noisy to make any meaningful inferences in the absence of task constraints. Indeed, it can be said that functional connectivity studies offer no way to separate signal from noise. This problem is compounded in psychedelic rsfMRI studies, as the black-box nature of the psychedelic experience makes it next to impossible to know what is going on in an individual's mind under the scanner. If psychedelics did nothing more than reduce the BOLD signals related to thought, while keeping the noise level constant, it would be enough to explain the findings of increased entropy and transition rate.
The paper's results were based on a paltry sample size of n=14 with less than 15mins of fMRI data per subject and a fairly low resolution of 3.4mm. The spatial maps generated were also low resolution as they were resampled from 2mm to 3.4mm. t-SNE is notably noisy, indeed the below figure shows the large variation in number of peaks generated from a single run.
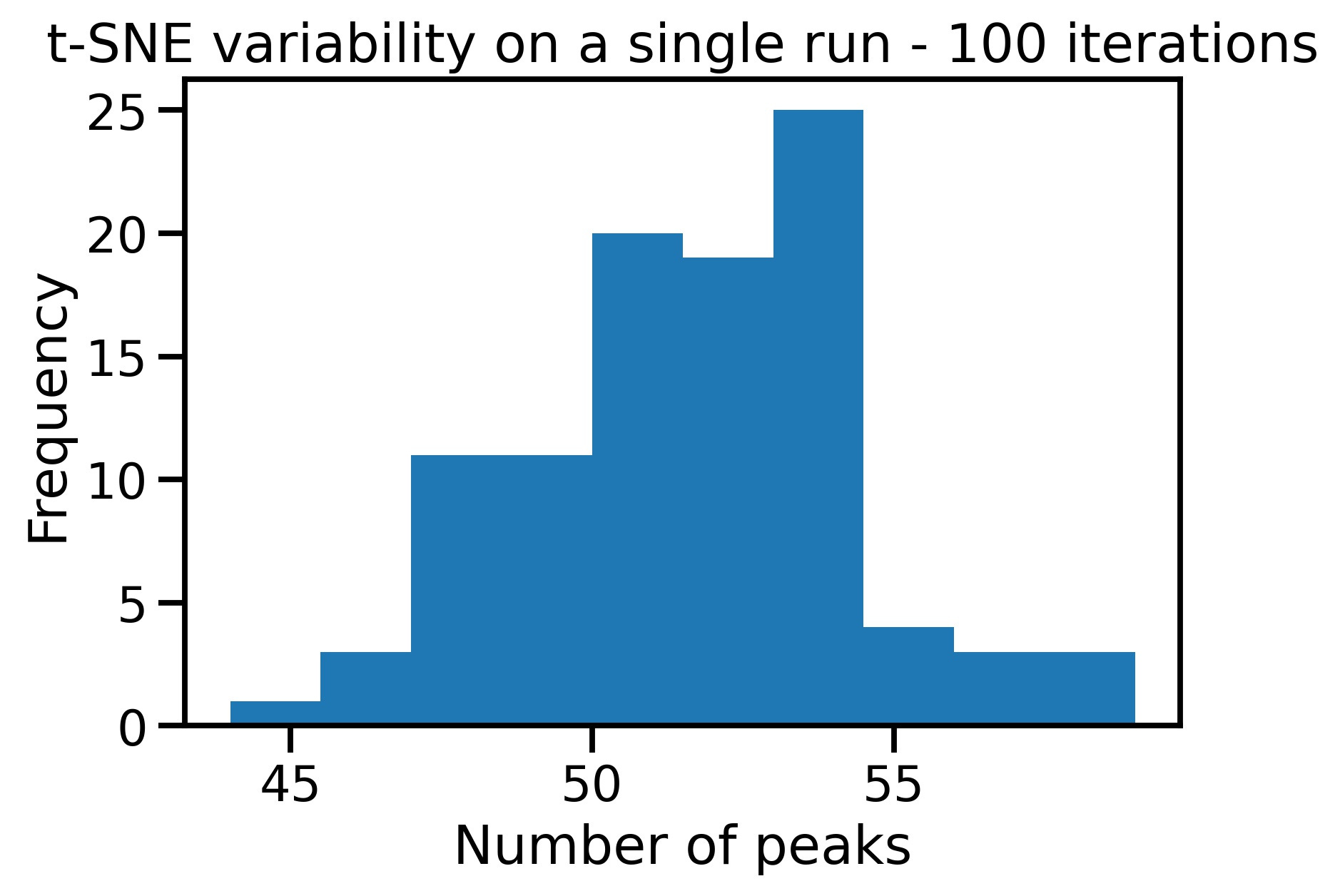
Open-Source LSD Data -
Robin Carhart-Harris et al. (2020). Neural correlates of the LSD experience revealed by multimodal neuroimaging. OpenNeuro. [Dataset] doi: 10.18112/openneuro.ds003059.v1.0.0 Setsompop K, Gagoski BA, Polimeni JR, Witzel T, Wedeen VJ, and Wald LL. (2012).
References
C.F. Beckmann, C.E. Mackay, N. Filippini, and S.M. Smith. Group comparison of resting-state FMRI data using multi-subject ICA and dual regression. OHBM, 2009. Neuroimage 80: 105-124.
Carhart-Harris, R. L., & Friston, K. (2019). REBUS and the anarchic brain: toward a unified model of the brain action of psychedelics. Pharmacological reviews, 71(3), 316-344.
Carhart-Harris, R. L., Leech, R., Hellyer, P. J., Shanahan, M., Feilding, A., Tagliazucchi, E., ... & Nutt, D. (2014). The entropic brain: a theory of conscious states informed by neuroimaging research with psychedelic drugs. Frontiers in human neuroscience, 8, 20.
David C. Van Essen, Stephen M. Smith, Deanna M. Barch, Timothy E.J. Behrens, Essa Yacoub, Kamil Ugurbil, for the WU-Minn HCP Consortium. (2013). The WU-Minn Human Connectome Project: An overview. NeuroImage 80(2013):62-79. **Ettinger, byJ. (2021, March 5). Compass pathways IPO brings $1 billion valuation. Psychedelic Spotlight. Retrieved December 11, 2021, from https://psychedelicspotlight.com/compass-pathways-ipo-brings-1-billion-valuation/. **Feinberg DA, Moeller S, Smith SM, Auerbach E, Ramanna S, Gunther M, Glasser MF, Miller KL, Ugurbil K, and Yacoub E. (2010). Multiplexed Echo Planar Imaging for sub-second whole brain fMRI and fast diffusion imaging.PLoS One 5:e15710. Fischl B. (2012). FreeSurfer. NeuroImage, 62:774-781.
Glasser, M. F., Sotiropoulos, S. N., Wilson, J. A., Coalson, T. S., Fischl, B., Andersson, J. L., ... & Wu-Minn HCP Consortium. (2013). The minimal preprocessing pipelines for the Human Connectome Project. Neuroimage, 80, 105-124.
Glennon, R. A., Titeler, M., & McKenney, J. D. (1984). Evidence for 5-HT2 involvement in the mechanism of action of hallucinogenic agents. Life sciences, 35(25), 2505-25
Gorgolewski K, Burns CD, Madison C, Clark D, Halchenko YO, Waskom ML, Ghosh SS. (2011). Nipype: a flexible, lightweight and extensible neuroimaging data processing framework in Python. Front. Neuroinform. 5:13.
Gorgolewski, K.J., Auer, T., Calhoun, V.D., Craddock, R.C., Das, S., Duff, E.P., Flandin, G., Ghosh, S.S., Glatard, T., Halchenko, Y.O., Handwerker, D.A., Hanke, M., Keator, D., Li, X., Michael, Z., Maumet, C., Nichols, B.N., Nichols, T.E., Pellman, J., Poline, J.-B., Rokem, A., Schaefer, G., Sochat, V., Triplett, W., Turner, J.A., Varoquaux, G., Poldrack, R.A. (2016). The brain imaging data structure, a format for organizing and describing outputs of neuroimaging experiments. Scientific Data, 3 (160044). doi:10.1038/sdata.2016.44
Griffiths, R. R., Richards, W. A., McCann, U., & Jesse, R. (2006). Psilocybin can occasion mystical-type experiences having substantial and sustained personal meaning and spiritual significance. Psychopharmacology, 187(3), 268-283 James, W. (1892).
The stream of consciousness. Psychology, 151-175. Jenkinson M, Bannister PR, Brady JM, and Smith SM. (2002).
Improved optimisation for the robust and accurate linear registration and motion correction of brain images. NeuroImage 17(2):825-841. Jenkinson M, Beckmann CF, Behrens TE, Woolrich MW, and Smith SM. (2012). FSL. NeuroImage, 62:782-790.
Lebedev, A. V., Lövdén, M., Rosenthal, G., Feilding, A., Nutt, D. J., & Carhart‐Harris, R. L. (2015). Finding the self by losing the self: Neural correlates of ego‐dissolution under psilocybin. Human brain mapping, 36(8), 3137-3153.
Lebedev, A. V., Kaelen, M., Lövdén, M., Nilsson, J., Feilding, A., Nutt, D. J., & Carhart‐Harris, R. L. (2016). LSD‐induced entropic brain activity predicts subsequent personality change. Human brain mapping, 37(9), 3203-3213.
Matthew F. Glasser, Stamatios N. Sotiropoulos, J. Anthony Wilson, Timothy S. Coalson, Bruce Fischl, Jesper L. Andersson, Junqian Xu, Saad Jbabdi, Matthew Webster, Jonathan R. Polimeni, David C. Van Essen, and Mark Jenkinson (2013).The minimal preprocessing pipelines for the Human Connectome Project
McCulloch, D. E., Knudsen, G. M., & Fisher, P. M. (2021). Psychedelic resting-state neuroimaging: a review and perspective on balancing replication and novel analyses. Milchenko M, and Marcus D. (2012).
Obscuring surface anatomy in volumetric imaging data. Neuroinformatics 2012 Sep 12 [Epub ahead of print]. Moeller S, Yacoub E, Olman CA, Auerbach E, Strupp J, Harel N, and Ugurbil K. (2010).
Multiband multislice GE-EPI at 7 tesla, with 16-fold acceleration using partial parallel imaging with application to high spatial and temporal whole-brain fMRI. Magn Reson Med. 63(5):1144-1153.
N. Filippini, B.J. MacIntosh, M.G. Hough, G.M. Goodwin, G.B. Frisoni, S.M. Smith, P.M. Matthews, C.F. Beckmann and C.E. Mackay. Distinct patterns of brain activity in young carriers of the APOE-ε4 allele. PNAS, 106(17):7209-14, 2009.
Petranker, R., Anderson, T., & Farb, N. (2020). Psychedelic research and the need for transparency: Polishing Alice’s Looking Glass. Frontiers in psychology, 11, 1681.
Blipped-controlled aliasing in parallel imaging for simultaneous multislice echo planar imaging with reduced g-factor penalty. Magn Reson Med. 67(5):1210-1224.
Singleton, S. P., Luppi, A. I., Carhart-Harris, R. L., Cruzat, J., Roseman, L., Deco, G., ... & Kuceyeski, A. (2021). LSD flattens the brain′ s energy landscape: evidence from receptor-informed network control theory. bioRxiv Steinhardt, Joanna, and Tehseen Noorani. 2020.
"Introduction: The Psychedelic Revival." Hot Spots, Fieldsights, July 21. https://culanth.org/fieldsights/introduction-the-psychedelic-revival
Van Essen, D. C., Smith, S. M., Barch, D. M., Behrens, T. E., Yacoub, E., Ugurbil, K., & Wu-Minn HCP Consortium. (2013). The WU-Minn human connectome project: an overview. Neuroimage, 80, 62-79.
Tseng, J., & Poppenk, J. (2020). Brain meta-state transitions demarcate thoughts across task contexts exposing the mental noise of trait neuroticism. Nature communications, 11(1), 1-12.
Van Essen, D. C., Ugurbil, K., Auerbach, E., Barch, D., Behrens, T. E., Bucholz, R., ... & WU-Minn HCP Consortium. (2012). The Human Connectome Project: a data acquisition perspective. Neuroimage, 62(4), 2222-2231.
Varley, T. F., Carhart-Harris, R., Roseman, L., Menon, D. K., & Stamatakis, E. A. (2020). Serotonergic psychedelics LSD & psilocybin increase the fractal dimension of cortical brain activity in spatial and temporal domains. Neuroimage, 220, 117049.
Xu J, Moeller S, Strupp J, Auerbach E, Feinberg DA, Ugurbil K, and Yacoub E. (2012). Highly accelerated whole brain imaging using aligned-blipped-controlled-aliasing multiband EPI. Proc. Int. Soc. Mag. Reson. Med. 20:2306.
Yarkoni, T., Markiewicz, C. J., de la Vega, A., Gorgolewski, K. J., Salo, T., Halchenko, Y. O., ... & Blair, R. (2019). PyBIDS: Python tools for BIDS datasets. Journal of open source software, 4(40).